Auditory & Vestibular Pathways
Abstract: Understanding the neurology of the auditory and vestibular systems allows the practitioner to challenge the function effectively. The auditory system has two parallel pathways of processing. The binaural sound localisation pathway and the monaural sound identification pathway. The vestibular system has five different receptors that measure the movement of the head in different planes. This then mediates vestibular reflexes which coordinate the bodies response to the heads movement.
Part 1: THE EAR
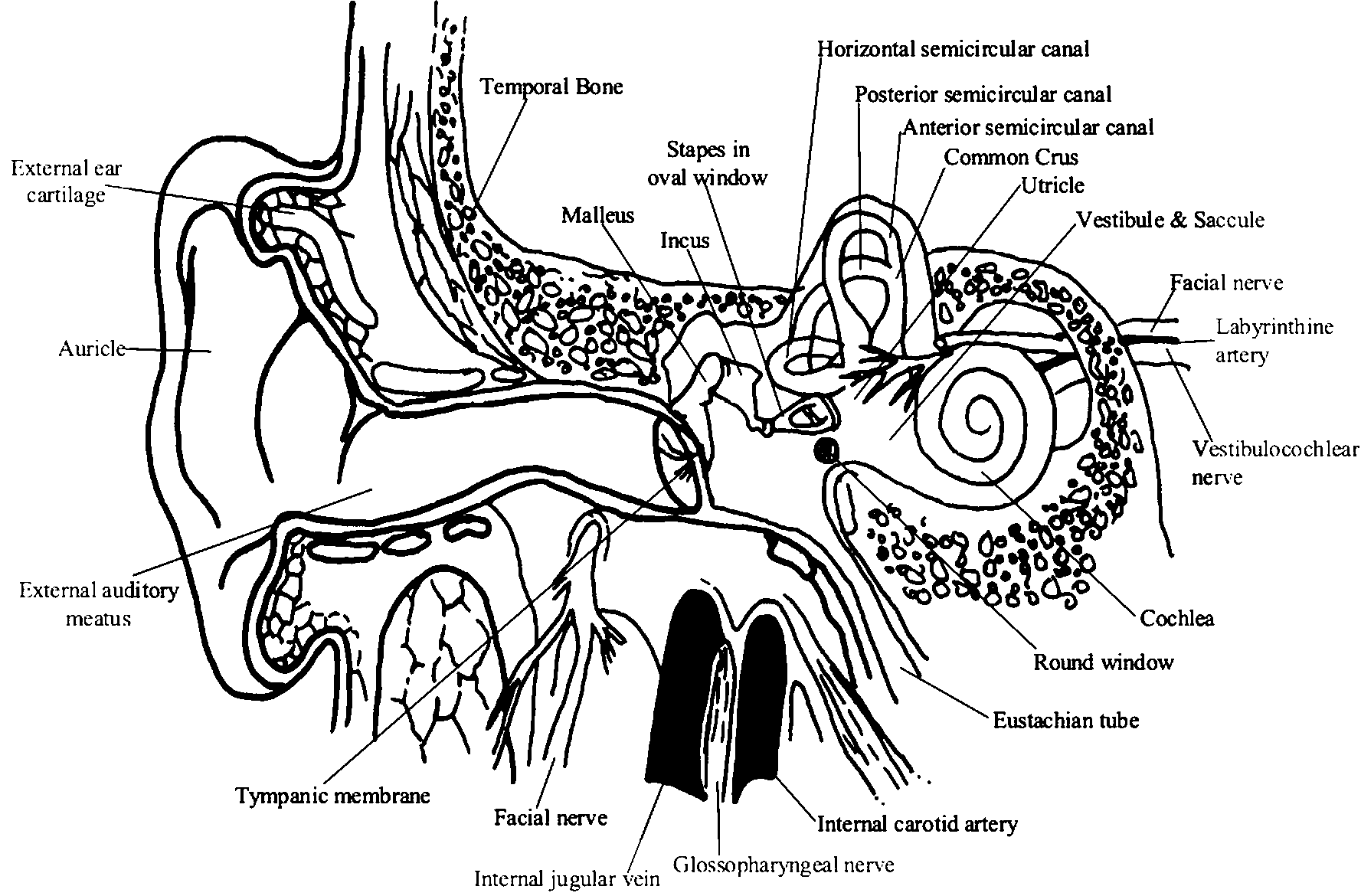
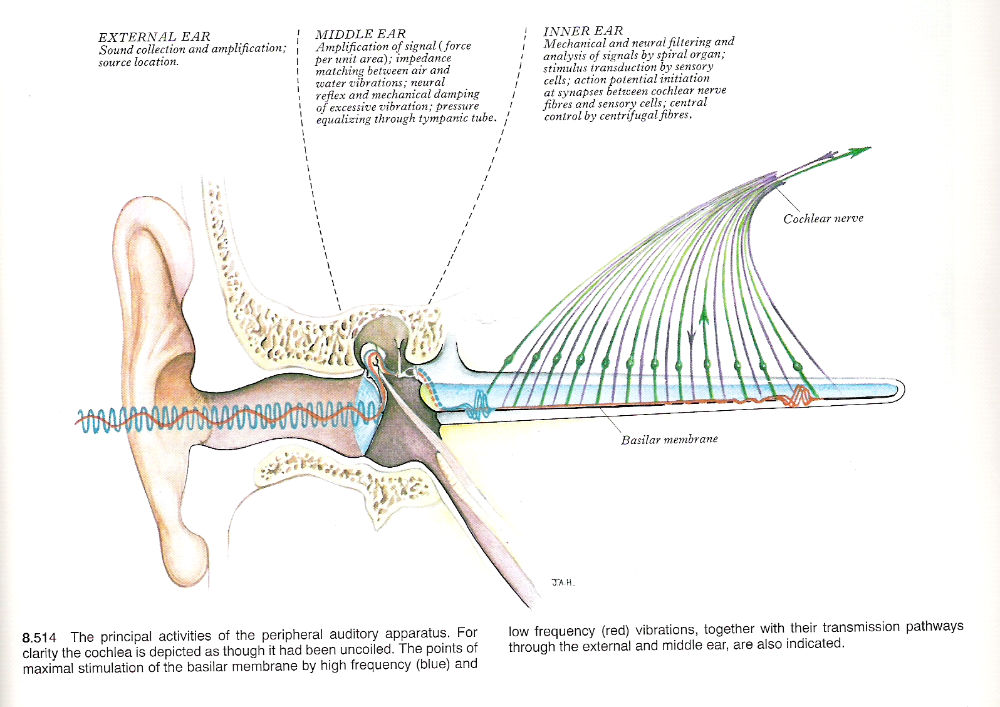
The peripheral auditory apparatus includes various parts of the ear, but of particular functional importance is the cochlear, which is a part of the membranous labyrinth. Each ear is a distance receptor for the collection, conduction, modification, amplification and analysis of complex waves of sound reaching it. These are transduced and coded into neural impulses in the afferent cochlear fibres of the vestibulocochlear nerve.
Sound waves consist of repetitive oscillations of the molecules in air, the waves vary in direction and distance from the source, intensity or energy content, mixture of frequencies in the train of waves and the phase of their vibrations.
The structural and functional design of ears makes them (within certain ranges) extremely sensitive to differences in frequency, intensity and phase. When ears are used binaurally, they are very effective range and direction finders. Ears are also very sensitive to the rate of change in all of these parameters. Frequency is expressed as cycles per second or Hertz (Hz). Young adult’s ears respond to frequencies of about 20 to 20,000 Hz.
-
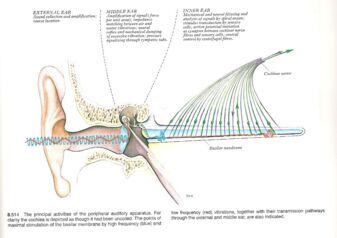
Fig 2: The Outer, Middle and Inner Ears, showing how different frequencies of sound are converted into neural signals. Source: Gray’s Anatomy -
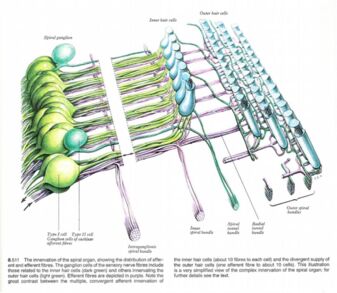
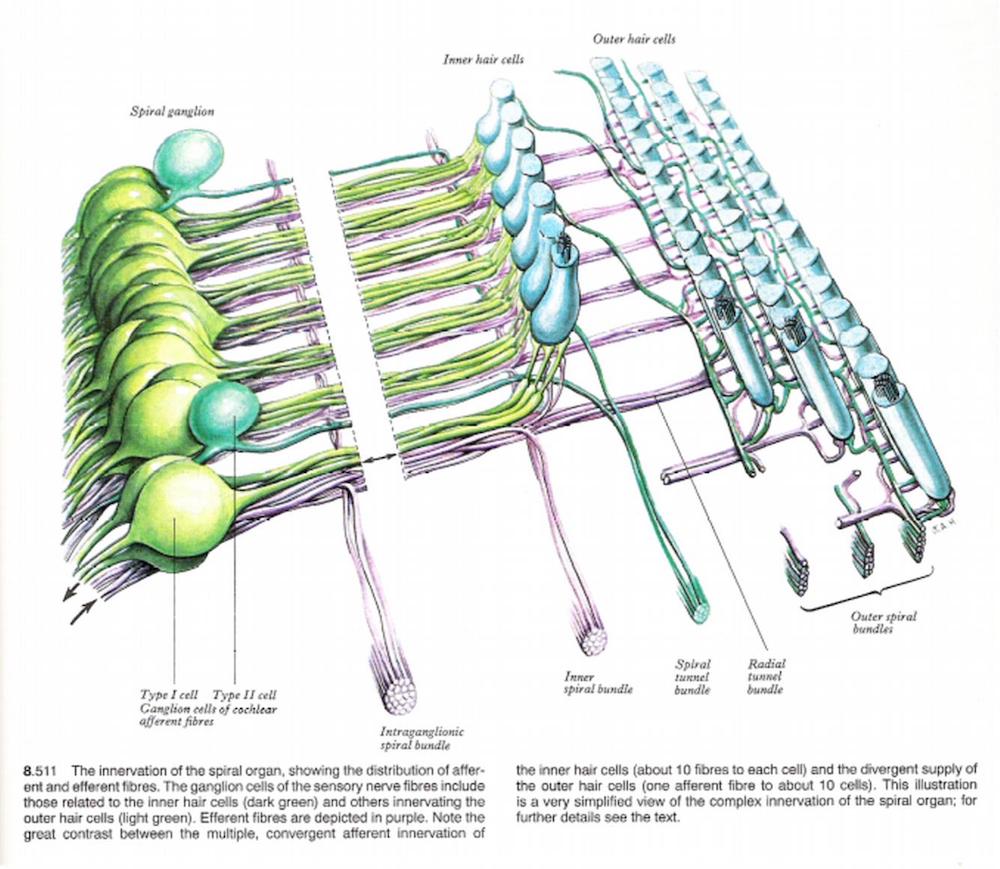
Type I cells have the following characteristics, 90% to 95% of spiral ganglion cells, they have radial branches that synapse with one or two inner hair cells, each inner hair cell has 20 or more type I cell fibres synapsing with it. Also type I cochlear nerve fibres respond to a narrow frequency range. This is because they are involved with sound identification
Type II cells have the following characteristics, they are widely distributed peripheral processes, they synapse with up to 10 outer hair cells, they are more sensitive to low intensity sounds than type I cells, they are less precisely tuned to frequency. They are involved with sound location.
Part 2: THE CENTRAL AUDITORY PATHWAYS
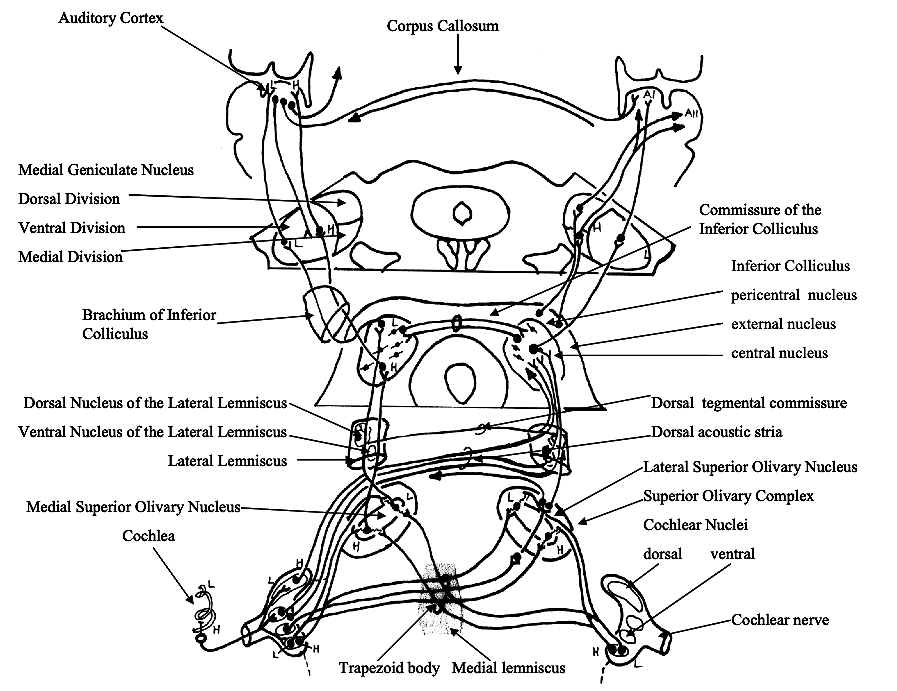
Part 3: LANGUAGE
Language can be defined as auditory stimuli, speech and linguistics. Auditory stimuli divided into, nonverbal auditory stimuli, auditory verbal stimuli - word lists, complex auditory verbal stimuli. Speech is divided into, automatic production, complex utterances.
People listening to noise or making simple loudness discrimination (nonverbal auditory stimuli) produces a bilateral cerebral hemispheric activation. This bilateral activation is mainly over the bilateral primary auditory cortex. When the acoustic and/or cognitive demands of the task were increased even slightly (eg. discrimination between two simple tones), the pattern of cerebral activation becomes more complex. Asymmetry of hemispheric activation is influenced by the cognitive strategy employed. Analytic strategy is more likely to increase left over right activation.
Listening to lists of words (auditory verbal stimuli) activates both cerebral hemispheres. Most studies demonstrate asymmetric activation favouring the left cerebral hemisphere. More complex auditory stimuli such as listening to short stories gives a similar cerebral activation pattern to the more simpler auditory verbal stimuli such as listening to word lists.
-
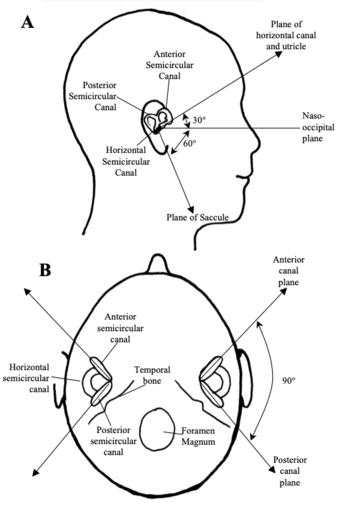
Fig 5: Orientation of the vestibular receptors. -
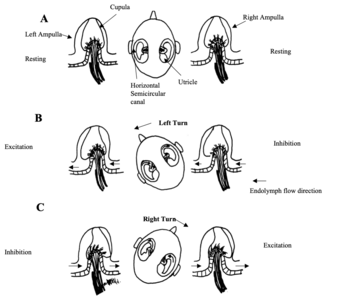
Fig 6: Response of the horizontal semicircular canals to head rotations in the horizontal plane. -
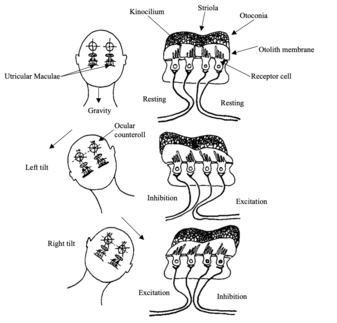
Fig 7: Responses of the utricular maculae to tilts of the head.
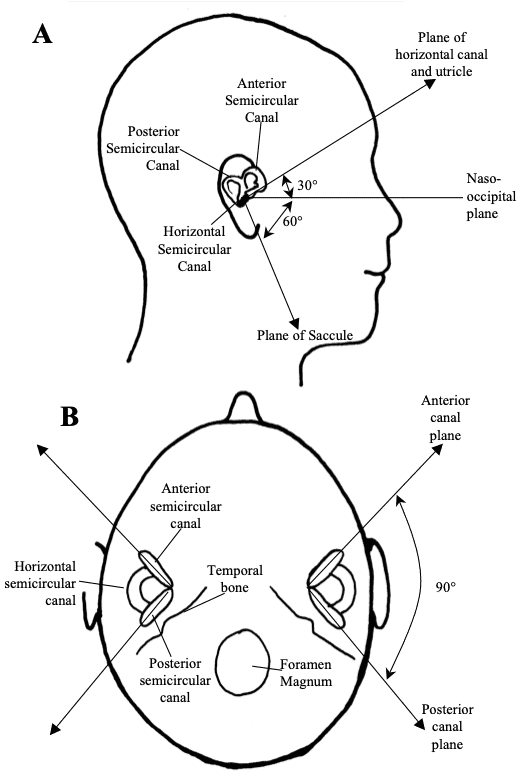
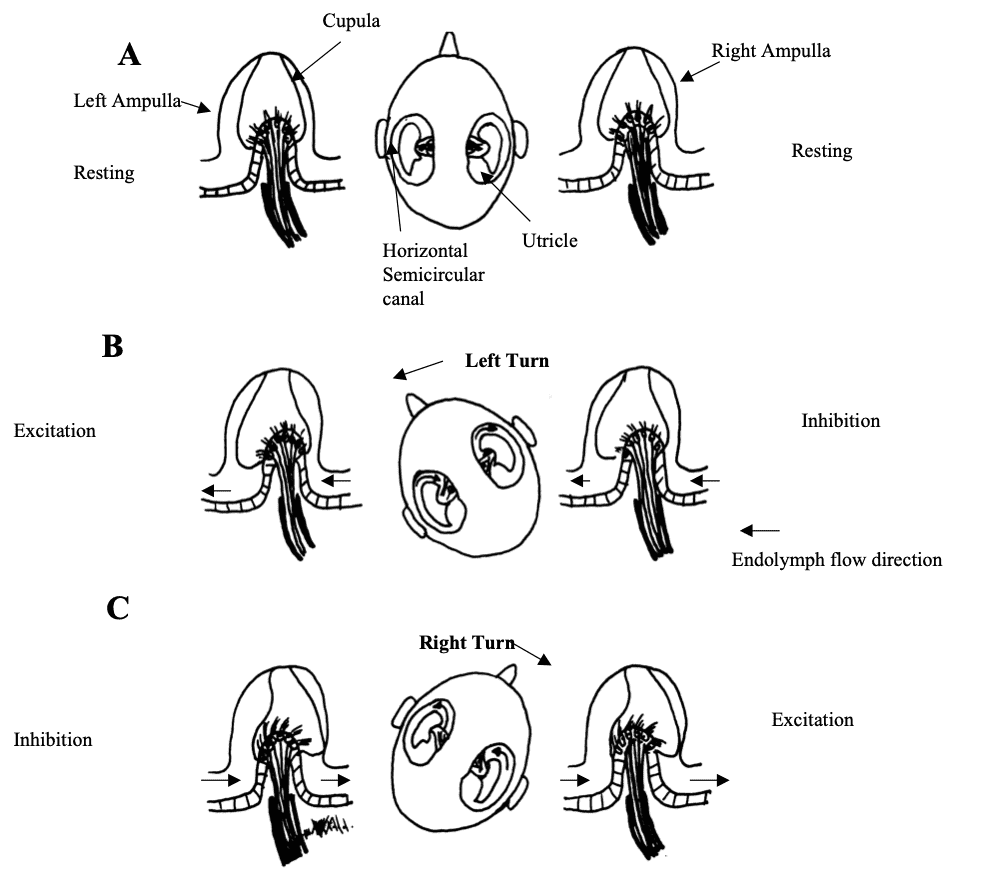
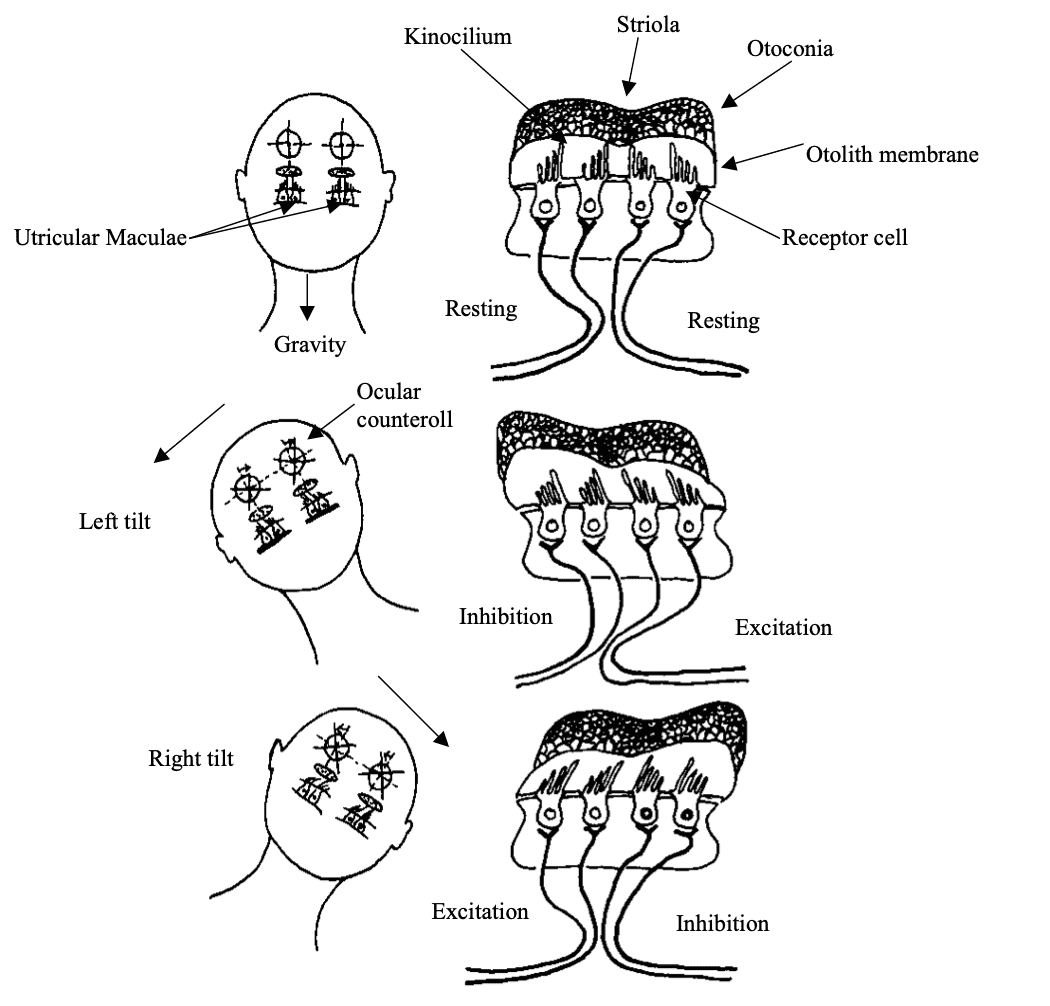
Fig 5: Orientation of the vestibular receptors. In the lateral view (A), the horizontal semicircular canal and the utricle lie in a plane that is tilted relative to the naso-occipital plane. In the axial view (B), the vertical semicircular canals lie at rightangles to each other. Source: Haines, Fundamental Neuroscience
Fig 6: Response of the horizontal semicircular canals to head rotations in the horizontal plane. (A) At rest the firing rate of horizontal canal afferents is equal on both sides. (B) is with a leftward turn and (C) is with a rightward head turn. There is receptor depolarisation (increase in firing) and afferent fibre excitation toward the side of the turn. There is receptor hyperpolarisation and afferent inhibition on the opposite side. Redrawn from Dickman 1997
Fig 7: Responses of the utricular maculae to tilts of the head. When the head is upright (A), the afferent fibres have equivalent firing rates on both sides of the striola. With leftward tilt (B) or rightward tilt (C), hair cells and their innervating afferents are either excited or inhibited, depending on their position relative to the striola; the weight of the otoconia causes the stereocilia to be deflected. Hair cells on the upslope side of the striola increase their firing rate, and those on the downslope side decrease their firing rate Redrawn from Dickman 1997
Part 4: THE VESTIBULAR SYSTEM
The vestibular system mediates the following motor activities, the ability to control posture, movement of the body relative to the external environment, and the movement of the eyes relative to the external environment. The vestibular system integrates peripheral sensory information from, the vestibular receptors, somatosensory receptors and visual receptors with motor information from the cerebellum and cerebral cortex.
The central processing of these inputs occurs rapidly. The output of the vestibular system is a signal to coordinate relevant muscle movements. The vestibular system is considered a special sense; most vestibular activity is conducted at a subconscious level. Situations that produce unusual vestibular stimulation, such as rough air in a plane or wave motion in ships produce acute vestibular perception. The possible resulting sensations are, dizziness, vertigo and nausea.
The vestibular system is an essential component in the formation of motor responses that are critical for daily function and survival. The vestibular system can be divided into five components.
- The peripheral receptor apparatus or the vestibular apparatus. This transduces head motion and position into neural information.
- The central vestibular nuclei. This is four nuclei in the brainstem that are responsible for receiving, integrating and distributing information that controls motor activities such as eye movements, head movements and postural reflexes.
- The vestibulo-ocular network arises from the vestibular nuclei and is involved in the control of eye movements.
- The vestibulospinal network coordinates head movements, axial musculature and postural reflexes.
- The vestibulo-thalamo-cortical network is responsible for the conscious perception of movement and spatial orientation.
The vestibular labyrinth consists of five separate receptor structures, three semicircular canals (horizontal canal, anterior canal, posterior canal) and two otolith organs (saccule and utricle). These five vestibular receptor organs in the inner ear complement each other in function. The semicircular canals transduce rotational head movements (angular accelerations). The otolith organs (utricle and saccule) respond to translational head movements (linear acceleration) or the orientation of the head relative to gravity. Each semicircular canal and otolith organ is spatially aligned so as to be most sensitive to movements in specific planes in 3-D space. In humans the horizontal semicircular canal and the utricle lie in a plane tilted approximately 30 above the horizontal. When a person walks or runs, the head is normally pitched downwards so the line of sight is a few metres in front of the feet. This orientation causes the plane of the horizontal canal and the utricle to be parallel with the earth horizontal and therefore perpendicular to gravity. The anterior and posterior semicircular canals and the saccule are arranged perpendicular to the plane of the horizontal canal and utricle. The anterior and posterior canals are arranged perpendicular to each other.
Part 5: THE VESTIBULAR REFLEXES
The Vestibular-ocular reflex allows fixating the eyes on an object of interest while the head is moving. For example reading a sign while walking down the street or in a moving vehicle. The vestibular system produces compensatory eye movements via a network of neural connections. The resulting stabilising eye movements are called the vestibular ocular reflex. These eye movements are equal in magnitude and in the opposite direction of head movement perceived by the vestibular system.
The vestibulo-ocular reflex occurs for any direction or speed of head movement which are rotational head movement, linear head movement, or a combination of both. The reflex can be suppressed at will if the observer wishes to focus on a moving target while the head moves in the same direction.
There are three types of rotational vestibule-ocular reflex eye movements, they are horizontal, torsional, and vertical movements. Each of the six pairs of eye muscles has to be controlled in unison to produce the required response.
Vertical eye movements are controlled by the vertical semicircular canals (Anterior & Posterior semicircular canals) and the saccule. Horizontal eye movements are controlled by the horizontal semicircular canals and the utricle. Torsional eye movements are controlled by the vertical semicircular canals (Anterior & Posterior semicircular canals) and the utricle.
There is also a linear vestibulo-ocular reflex. This involves linear head movement and depends on input from the Otolith organ. Vertical displacements of the body such as walking, running or travelling in a car produce oppositely directed vertical eye movements to stabilise the gaze.
When head rotations are large, for example a 360° body turn, compensatory eye movements take another form. The vestibulo-ocular reflex directs eyes slowly in the direction opposite to head motion. This is called slow phase. When the eye reaches the limit of how far it can turn in the orbit, it springs back rapidly to a central position, moving the same direction as the head. This is called fast phase. Another slow phase begins. The combination of slow phase and fast phase is called Nystagmus
The vestibulospinal reflex influences muscle tone, produces reflex postural adjustments of the head, produces reflex postural adjustments of the body.
The cognitive perception of motion and spatial orientation happen at the thalamocortical level. This comes from a convergence of information from the vestibular, visual and somatosensory systems. This is the vestibulo-thalamo-cortical network.
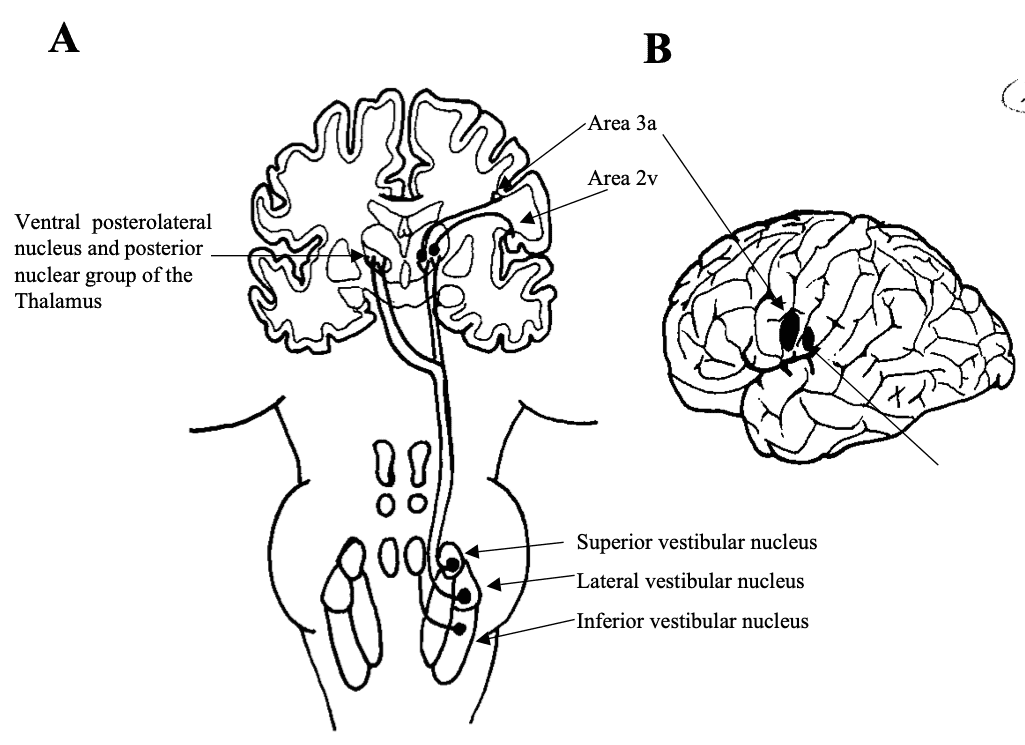
Electrical stimulation of the vestibulo-thalamic areas produces sensation of movement and dizziness. The two cortical areas respond to vestibular stimulation are areas 2v and 3a. Electrical stimulation of 2v neurones produces sensations of moving, spinning and dizziness. 2v neurones respond to head movements and receive thalamic projections from the posterior nuclear group. 2v neurones also receive visual and proprioceptive information. 2v is probably involved with motion perception and spatial orientation, because it has reciprocal connections with other parietal areas involved in similar functions (ie. Areas 5 and 7).
3a receives inputs from the somatosensory system. These cells project to area 4 of the motor cortex. This area is thought to integrate motor control of the head and body.
|
Receptor |
Ocular movements |
|
Horizontal semicircular canal |
Horizontal eye movements |
|
Anterior semicircular canal |
Vertical and torsional eye movements |
|
Posterior semicircular canal |
Vertical and torsional eye movements |
|
Utricle |
Horizontal and torsional eye movements |
|
Saccule |
Vertical eye movements, linear head movements |
-
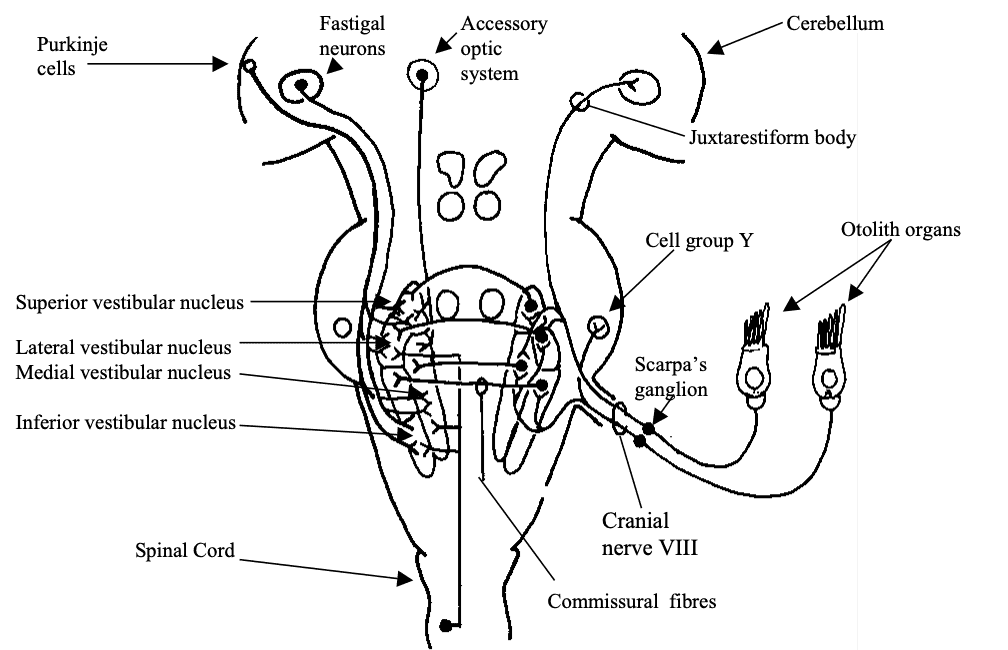
Fig 8: Afferents to the Vestibular nuclei. Open cell bodies represent inhibitory projections. Redrawn from Dickman 1997 -
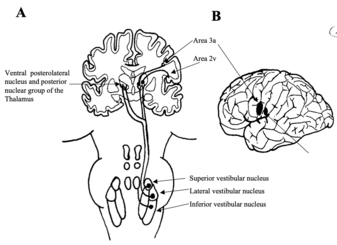
Fig 9: The vestibulo-thalamo-cortical pathway. Vestibular input arises from the vestibular nuclei as vestibulothalamic fibres and is relayed to the cortex as thalamocortical fibres (A). Areas 3a and 2v (B) are the main cortical regions that receive this input. Redrawn from Dickman 1997